ARTÍCULOS Y PUBLICACIONES
MOLECULAS DE SEÑALIZACIÓN DEL EPITELIO BUCAL: EVALUACIÓN DEL EFECTO DEL CAMPO DE INFORMACIÓN EN EL CÁNCER DE MAMA
S. S. KONOVALOV, Doctor en Medicina, Profesor Catedrático, kvantdnk@yandex.ru,
Director del Instituto de Medicina Informativa de la MANEB (Academia Internacional de Ciencias
de la Ecología, de la Seguridad del Hombre y de la Naturaleza), San Petersburgo.
S.S. KONOVALOV, Doctor of Medicine, Professor, kvantdnk@yandex.ru,
Director of Institute of Information Medicine MANEB, St. Petersburg.
Uno de los métodos más comunes y efectivos de diagnóstico molecular de enfermedades es la inmunohistoquímica. Sin embargo, su valor pronóstico es en gran medida limitado debido a la invasividad del acceso para obtener el material estudiado. En este sentido, la tarea urgente de la medicina molecular moderna es la búsqueda de objetos de investigación celular y tisular, cuyo recibo esté disponible de forma no invasiva. En los últimos años, el epitelio bucal (EB) ha atraído la atención de los investigadores como posible material para diagnósticos no invasivos.
El muestreo del material desde la superficie interna de la mejilla es un procedimiento no invasivo, además, este objeto de investigación es muy informativo (estudia la expresión de moléculas de señalización, la capacidad de evaluar las características electrocinéticas de las células, etc.) y puede usarse para el diagnóstico intravital de enfermedades socialmente significativas.
El EB puede considerarse como una zona fronteriza entre el entorno externo e interno del cuerpo. Por lo tanto, los cambios en la actividad funcional de las células BE (procesos de renovación y diferenciación celular, expresión de diversas moléculas de señalización) reflejan en gran medida el estado de la homeostasis local y sistémica del cuerpo o su alteración en condiciones patológicas.
Por lo tanto, el EB disponible para examen histológico intravital puede servir como una fuente de información diagnóstica y pronóstica importante sobre el estado de salud, el estrés, la influencia de factores ambientales, la patología somática y la edad biológica de una persona.
Se realizó un estudio sobre la expresión de una serie de moléculas de señalización clave que llevan a cabo interacciones intercelulares neuroinmunoendocrinas, que pueden servir como marcadores de procesos homeostáticos y de adaptación compensatoria en el cuerpo humano: proteínas transcripcionales, moléculas de adhesión, grupos de diferenciación de células inmunocompetentes residentes y proteínas involucradas en la regulación del ciclo celular.
El objetivo del estudio fue analizar el nivel de expresión de las moléculas de señalización en el epitelio bucal en pacientes con cáncer de mama para evaluar la efectividad del campo de información.
Para el análisis de la expresión de 21 moléculas de señalización (MITF, BMP, SMAD2, ERBB3, WNT5A, RON, CD90, CD73, CD51, CD64, CD105, CXCL12, Pax6, Chx10, RTF10, FOXA2, Oct2, PROX1, Nkx2.5, MAB045, grelina) las células se obtuvieron usando un cepillo citológico de la membrana mucosa de la mejilla (células epiteliales, neutrófilos, linfocitos, macrófagos). Se tomó una muestra bucal de 50 pacientes de 37 a 73 años.
Todos los pacientes se dividieron en 3 grupos: grupo 1 - mujeres sin patología oncológica (a - antes de la sesión de exposición al efecto informativo energético, b - después de la sesión de exposición al efecto informativo energético), grupo 2 - mujeres con cáncer de mama que no asisten a las sesiones y recibiendo solo terapia estándar, grupo 3 - mujeres con cáncer de mama (a - antes de una sesión de exposición al efecto informativo energético, b - después de una sesión de exposición al efecto informativo energético).
Para llevar a cabo una reacción inmunohistoquímica con anticuerpos contra las moléculas de señalización estudiadas, se usó un protocolo estándar de un solo paso con el desenmascarado de antígeno a alta temperatura en el tampón de citrato (pH = 6,0). Se usó un kit universal que contenía inmunoglobulinas anti-ratón biotiniladas (Dako) como anticuerpos secundarios. Para visualizar la reacción, se usó un complejo de avidina con peroxidasa de rábano picante biotinilada, luego se usó diaminobencidina (kit ABC, Dako) para la manifestación.
El estudio morfométrico se llevó a cabo utilizando un sistema de análisis de imagen microscópico computarizado que consta de un microscopio Nikon Eclipse E400, una cámara digital Nikon DXM1200, una computadora personal basada en Intel Pentium 4 y el software “Video Test - Morfology 5.2.”. En cada caso, se analizaron 5 campos de visión con un aumento de x400. El área de expresión de estos marcadores se calculó como la relación del área ocupada por las células inmunopositivas al área total de células en el campo de visión y se expresó como un porcentaje.
Para el procesamiento de datos estadísticos, se usó la prueba de Wilcoxon para comparar varias muestras.
1.1 Influencia del efecto del campo de información sobre la expresión de moléculas: grupos de diferenciación de células inmunocompetentes en las pacientes con cáncer de mama
Moléculas: los grupos de diferenciación (cluster of differentiation) son antígenos diferenciadores de leucocitos humanos. Esta clasificación se propuso en 1982 para la identificación y el estudio de proteínas de membrana de superficie de leucocitos. Los antígenos CD (o de otro modo marcadores CD) pueden ser proteínas que sirven como receptores o ligandos que participan en la interacción de las células entre sí y son componentes de una cascada de rutas de señalización específicas. Sin embargo, pueden ser proteínas que realizan otras funciones (por ejemplo, proteínas de adhesión celular).
1.1.1. Expresión del marcador CD51
El marcador CD51 (integrina alpha-V) es una glucoproteína integrina y un producto de ITGAV. La integrina alpha-V es un receptor para vitronectina, citotactina, fibronectina, fibrinógeno, laminina, metaloproteasa de matriz 2, osteopontina, osteomodulina, protrombina, trombospondina y factor von Willebrand. El CD51 reconoce la secuencia de aminoácidos específica de glicina-prolina-arginina (R-G-D) en un amplia gama de ligandos. La integrina alpha-V consiste en una cadena pesada y ligera unida por un enlace disulfuro. La integrina alpha-V forma un diámetro con integrinas beta-1, beta-3, beta-5, beta-6 o beta-8. Por lo tanto, el marcador CD51 refleja varios aspectos de las interacciones intercelulares.
Se encontró que en pacientes sin patología oncológica después de la exposición al efecto informativo energético, el área de expresión del marcador CD51 aumentó en un 30% en comparación con el mismo indicador antes de la exposición (Fig. 1). En pacientes con cáncer de mama en los grupos de terapia estándar y antes de la exposición al efecto informativo energético, el área de expresión de CD51 fue casi 2 veces menor en comparación con el grupo de mujeres sanas antes de la exposición (Fig. 1). Durante la exposición al efecto informativo energético, el área de expresión del marcador CD51y de pacientes con cáncer de seno aumentó 2 veces y alcanzó el valor en el grupo 1 (Fig. 1).
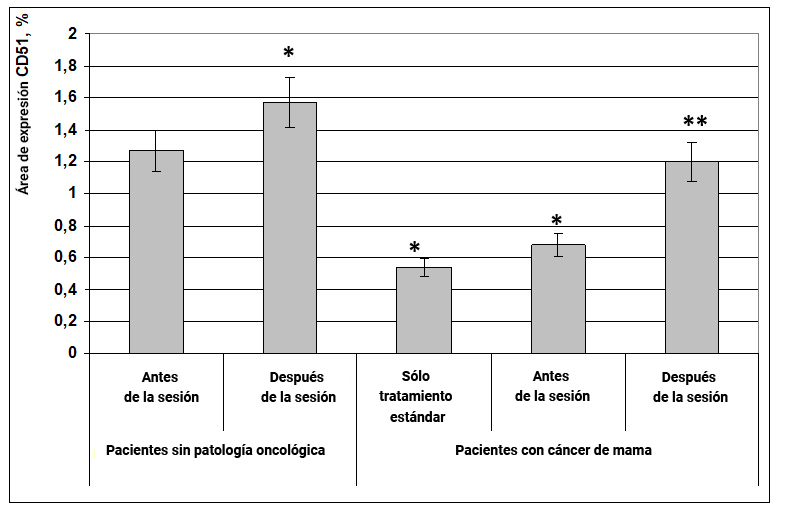
Fig. 1. Influencia del efecto informativo energético sobre la expresión del marcador CD51 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
Por lo tanto, en el cáncer de mama, hay una disminución en la expresión de la glucoproteína CD51 en las células del frotis bucal, y el efecto informativo energético ayuda a restaurar este parámetro a la normalidad.
1.1.2. Expresión del marcador CD64
El marcador CD 64 es un factor desencadenante clave en los mecanismos de la citólisis de células tumorales por monocitos humanos y neutrófilos activados por interferón. Esta glucoproteína juega un papel central en la citotoxicidad celular dependiente de anticuerpos.
En pacientes sin patología oncológica, después de la exposición al efecto informativo energético, el área de expresión del marcador CD64 aumentó en un 25% en comparación con el mismo indicador antes de la exposición (Fig. 2). En pacientes con cáncer de mama con terapia estándar y antes de la exposición al efecto informativo energético, el área de expresión de CD64 fue 75% menor en comparación con el grupo de mujeres sanas antes de la exposición (Fig. 2, 3). Cabe señalar que después de la exposición al efecto informativo energético, el área de expresión del marcador CD64 aumentó 2 veces y alcanzó el valor en el grupo de mujeres sanas (Fig. 1).
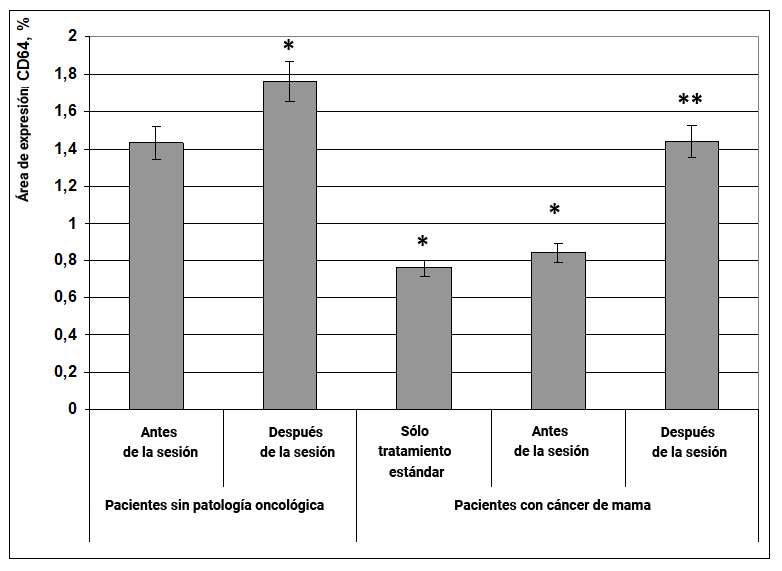
Fig. 2. Influencia del efecto informativo energético sobre la expresión del marcador CD64 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
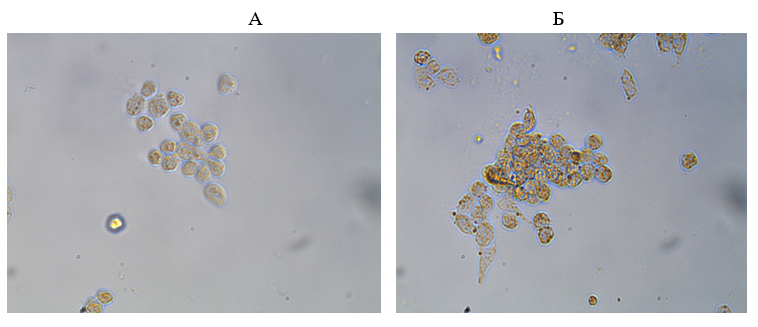
Fig. 3. Expresión del marcador CD64 en pacientes con cáncer de mama: A - antes de exponerse al efecto informativo energético, B - después de exponerse al efecto informativo energético, inmunohistoquímica, x200.
Por lo tanto, la influencia positiva del efecto informativo energético sobre las pacientes con cáncer de mama se asocia con la activación de la proteína anticancerogénica CD64 y la inmunidad celular.
1.1.3. Expresión del marcador CD73
Se sabe que la glucoproteína CD73 activa la proliferación de linfocitos y su adhesión al endotelio vascular. Una disminución en la expresión del marcador CD73 es la causa del desarrollo de inmunodeficiencias, que pueden correlacionarse con una disminución en la protección anticancerígena y un aumento en la incidencia de tumores.
Se demostró que en pacientes con cáncer de mama con terapia estándar y antes de la exposición al efecto informativo energético, el área de expresión de CD73 era en un 27% menor que en el grupo de mujeres sanas antes de la exposición (Fig. 4). Bajo la influencia del efecto informativo energético, el área de expresión del marcador CD73 en las células en pacientes con cáncer de mama tendió a aumentar, pero este resultado no fue confiable. En mujeres sin patología oncológica, después de la exposición tampoco se observaron cambios en la expresión del marcador estudiado. (Fig. 4).
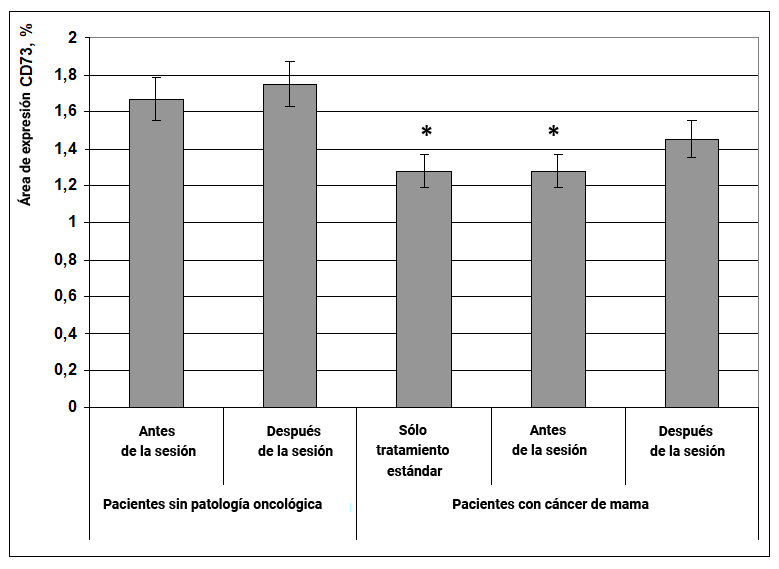
Fig. 4. Influencia del efecto informativo energético sobre la expresión del marcador CD73 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
Es probable que el efecto informativo energético no haya afectado la expresión del marcador CD73 en pacientes con cáncer de mama debido al hecho de que este indicador solo se redujo ligeramente en comparación con el grupo de mujeres sanas.
1.1.4. Expresión del marcador CD90
El marcador CD90 es un marcador de células madre y contribuye a su transición a formas comprometidas, especialmente al regular la diferenciación de células de origen neuronal y estromal.
La glucoproteína CD90 estimula la formación de neuronas cerebrales altamente diferenciadas, participando en la formación de la función cognitiva. Regula la función de los canales de calcio, el crecimiento del axón y la transmisión sináptica. Este marcador es un factor antiapoptótico en relación con las células linfoides, especialmente los linfocitos T.
Una disminución de su expresión en una población de neuronas o células linfoides implica el desarrollo de tumores neurogénicos y linfoblastosis. Una disminución de su expresión en los endoteliocitos conduce a una adhesión endotelial deteriorada y a la ruptura del vaso y hemorragia.
En pacientes sin patología oncológica, el efecto informativo energético no afectó la expresión de la glucoproteína CD90 antitumoral, pero en pacientes con cáncer de mama este efecto causó un impacto significativo. Por lo tanto, en pacientes con neoplasias, el área de expresión del marcador CD90 en comparación con mujeres sanas disminuyó en un 45% (Fig. 5). Bajo la influencia del campo energético de información, el área de expresión de CD90 aumentó en un 50% (Fig. 5) y alcanzó los valores típicos para mujeres sanas.
La restauración del nivel de expresión del marcador antitumoral CD90 bajo la influencia del campo energético de información indica un aumento en la función inmune del cuerpo, lo que puede conducir a una desaceleración en el crecimiento del tumor.
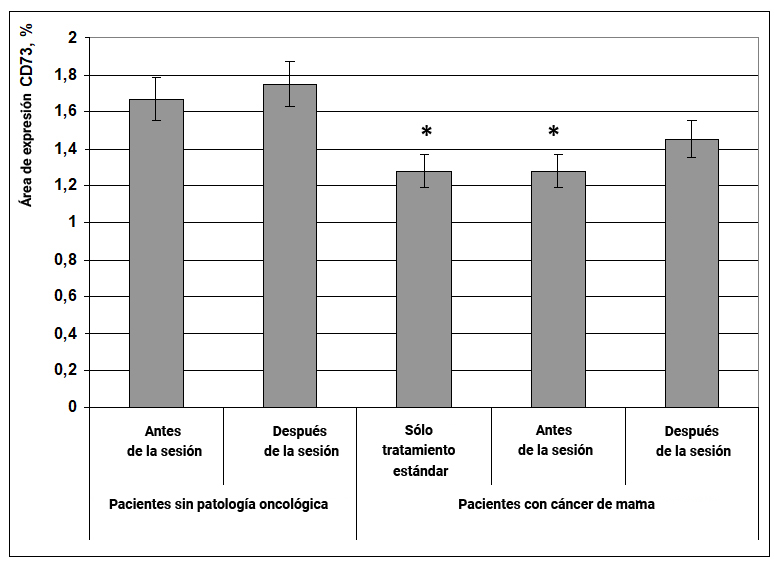
Fig. 5. Influencia del efecto informativo energético sobre la expresión del marcador CD90 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
1.1.5. Expresión del marcador CD105
La molécula CD105 (endoglina) activa las funciones de macrófagos, fibroblastos, células de músculo liso y células endoteliales.
La glucoproteína CD105 participa en el mantenimiento de la estructura del citoesqueleto de las células y afecta su migración. El marcador CD105 juega un papel importante en el desarrollo del sistema cardiovascular y la remodelación vascular. Las mutaciones en el gen que codifica la proteína CD105 están involucradas en el desarrollo de una enfermedad autosómica dominante conocida como telangiectasia hemorrágica hereditaria tipo 1, que se manifiesta por malformaciones arteriovenosas de los órganos.
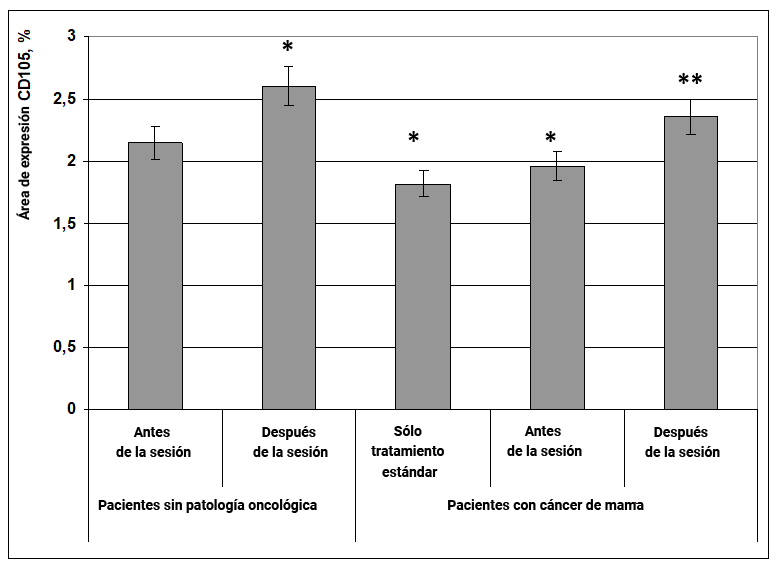
Fig. 6. Influencia del efecto informativo energético sobre la expresión del marcador CD105 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
Se encontró que en pacientes sin patología oncológica después de la exposición al efecto informativo energético, el área de expresión del marcador CD105 aumentó con certitud en un 22% en comparación con el mismo indicador antes de la exposición (Fig. 6). En pacientes con cáncer de mama en los grupos de terapia estándar y antes de que se realizara la exposición al efecto informativo energético, el área de expresión de CD105 era un 37% menor en comparación con el grupo de mujeres sanas antes de la exposición (Fig. 6). Durante el impacto informativo y energético, el área de expresión del marcador CD105 en pacientes con patología oncológica aumentó a los valores obtenidos en mujeres sanas (Fig. 6).
Por lo tanto, la información y el efecto informativo energético mejoran la expresión del marcador CD105 en mujeres sanas y pacientes con cáncer de mama, lo que indica su capacidad para activar macrófagos tisulares y normalizar la función vascular.
1.2. Influencia del efecto del campo de información sobre la expresión de factores
de transcripción y diferenciación celular en las pacientes con cáncer de mama
Los factores de transcripción son proteínas que controlan la transferencia de información de una molécula de ADN a la estructura del ARNm (transcripción) uniéndose a sitios de ADN específicos. Los factores de transcripción realizan su función independientemente o en combinación con otras proteínas. Proporcionan una disminución (represores) o un aumento (activadores) de la constante de unión de la ARN polimerasa a las secuencias reguladoras del gen regulado.
Una característica definitoria de los factores de transcripción es la presencia en su composición de uno o más dominios de unión al ADN que interactúan con regiones características de ADN ubicadas en las regiones reguladoras de los genes. Otras proteínas que juegan un papel clave en la regulación de la expresión génica, como los coactivadores, histonas acetilasas, quinasas, metilasas, no tienen dominios de unión al ADN y, por lo tanto, no pueden asignarse a factores de transcripción.
Los factores de transcripción son necesarios para la regulación de la expresión génica y se encuentran en todos los organismos vivos. Su número, tanto absoluto como específico, aumenta con el aumento del tamaño del genoma. Se han encontrado más de 2.600 proteínas con un dominio de unión al ADN en el genoma humano, y se cree que la mayoría de ellas son factores de transcripción. Por lo tanto, aproximadamente el 10% de todos los genes en el genoma están codificados por factores de transcripción. Por lo tanto, son la familia más grande de proteínas humanas. Además, muchos genes están regulados por la interacción corporativa de una gran cantidad de factores de transcripción diferentes, lo que permite que cada uno de los genes proporcione una forma única de regulación en el proceso de desarrollo del organismo.
La función de los factores de transcripción es garantizar la lectura e interpretación de la información genética. Se unen al ADN y ayudan a iniciar un programa para aumentar o disminuir la transcripción génica. Por lo tanto, son vitales para el funcionamiento normal del cuerpo en todos los niveles. Los procesos más importantes en los que intervienen factores de transcripción son: regulación de la expresión génica basal, regulación de la ontogénesis, respuesta a señales intracelulares e influencias ambientales.
La actividad transcripcional de fondo es proporcionada por un conjunto de factores de transcripción comunes a todos los genes. Una clase importante de factores de transcripción eucariotas son los GTF (factores de transcripción generales). Muchos de sus representantes no se unen al ADN directamente, sino que forman parte del complejo de iniciación de la transcripción (complejo preiniciador), que interactúa directamente con la ARN polimerasa. Los GTF más comunes son TFIIA, TFIIB, TFIID (asociados con el denominado cuadro TATA (elemento promotor)), TFIIE, TFIIF y TFIIH. Además de los factores de transcripción necesarios para la expresión de todos los genes, también hay factores de transcripción específicos que permiten activar / desactivar ciertos genes en el momento adecuado.
Muchos factores de transcripción de organismos multicelulares están involucrados en asegurar su desarrollo. Actuando de acuerdo con el programa genético y / o en respuesta a influencias externas, inician o suprimen la transcripción de ciertos genes, lo que implica cambios en la morfología celular, diferenciación celular, morfogénesis, organogénesis, etc. Por ejemplo, la familia de factores de transcripción homeobox es crítica. para la formación de la morfología corporal correcta en organismos desde Drosophila hasta humanos. Las mutaciones en los genes de estas proteínas (mutaciones de la homeosis) en Drosophila conducen a trastornos graves en la diferenciación de los segmentos del cuerpo de estos insectos (por ejemplo, desarrollo de patas en lugar de antenas).
Otro ejemplo de este grupo de factores de transcripción es el gen de la región determinante del sexo Y (SRY, Sex-determing Region Y), que desempeña un papel importante en la determinación del sexo de una persona.
La regulación coordinada de la interacción de las células de un organismo multicelular se lleva a cabo mediante la liberación de moléculas especiales (hormonas, citocinas, etc.) que causan una cascada de señalización en las células objetivo. Si la señal causa un cambio en el nivel de expresión de ciertos genes, los factores de transcripción son a menudo el eslabón final en la cascada. La vía de señalización de estrógenos es un ejemplo de una cascada corta que incluye un factor de transcripción del receptor de estrógenos: el estrógeno es secretado por los tejidos de la placenta y el ovario, atraviesa la membrana plasmática de las células receptoras y se une a su receptor en el citoplasma. El receptor de estrógenos ingresa al núcleo y se une a un sitio de ADN específico, cambiando la regulación de la transcripción del gen correspondiente.
Los factores de transcripción no son los únicos enlaces finales de las cascadas de señalización que surgen en respuesta a diversos estímulos externos, sino que también pueden ser efectores en las cascadas de señalización inducidas por las influencias ambientales. Por ejemplo, el factor de choque térmico (HSF) activa los genes responsables de la supervivencia a altas temperaturas (como las chaperonas), el factor inducible por hipoxia (HIF), al tiempo que reduce la concentración de oxígeno, la SREBP (sterol regulatory element binding protein - proteína de unión al elemento regulador de esteroles) ayuda a mantener el contenido requerido de lípidos en las células.
Muchos factores de transcripción, especialmente oncógenos y supresores de cáncer, están involucrados en la regulación del ciclo celular. Determinan la transición de una fase del ciclo celular a otra, la frecuencia de las divisiones y la intensidad del crecimiento. Uno de los factores de transcripción más conocidos es el oncógeno Myc, que desempeña un papel importante en el crecimiento celular y su apoptosis.
En los organismos eucariotas, los procesos de transcripción y traducción están separados espacialmente, se producen en el núcleo y el citoplasma, respectivamente. Después de la síntesis, los factores de transcripción deben penetrar en el núcleo, rompiendo la doble membrana. Muchas proteínas que funcionan en el núcleo tienen una señal de localización nuclear, una región específica de la cadena de polipéptidos que se dirige a la proteína en el núcleo. Para muchos factores de transcripción, la translocación es un factor clave en la regulación de su actividad. Las clases importantes de factores de transcripción, como algunos receptores nucleares, deben unirse primero al ligando en el citoplasma y solo luego transportarse al núcleo.
En los eucariotas, los genes que no se transcriben constantemente a menudo se encuentran en heterocromatina (regiones de ADN estrechamente empaquetadas por la unión de histonas y organizadas en fibrillas de cromatina compactas). El ADN en heterocromatina no está disponible para muchos factores de transcripción. Para que los factores de transcripción se unan al ADN, la heterocromatina debe transformarse en eucromatina, generalmente a través de modificaciones de histonas. El sitio de unión de los factores de transcripción en el ADN puede no ser accesible si está unido por otro factor de transcripción. Los pares de factores de transcripción pueden desempeñar un papel antagónico (activador - represor) en la regulación de la actividad de un gen.
1.2.1 Expresión del factor de transcripción Pax6
El factor de transcripción Pax6 juega un papel clave en el desarrollo de varios órganos. La expresión de este factor determina la intensidad de la proliferación celular y el número de poblaciones celulares, regulando la relación entre la reproducción celular y su muerte (apoptosis). El efecto regulador más importante de Pax6 es el desarrollo del páncreas, el tejido nervioso y los órganos sensoriales, especialmente la retina, donde determina la formación de capas retinianas y la polaridad de los fotorreceptores.
Se encontró que en pacientes sin cáncer, la expresión del factor de transcripción Pax6 fue en un 23% menor en comparación con este indicador en mujeres con cáncer de mama. El efecto informativo energético contribuyó a la restauración de la expresión del marcador Pax6 a los valores característicos de las mujeres sin patología oncológica (Fig. 7).
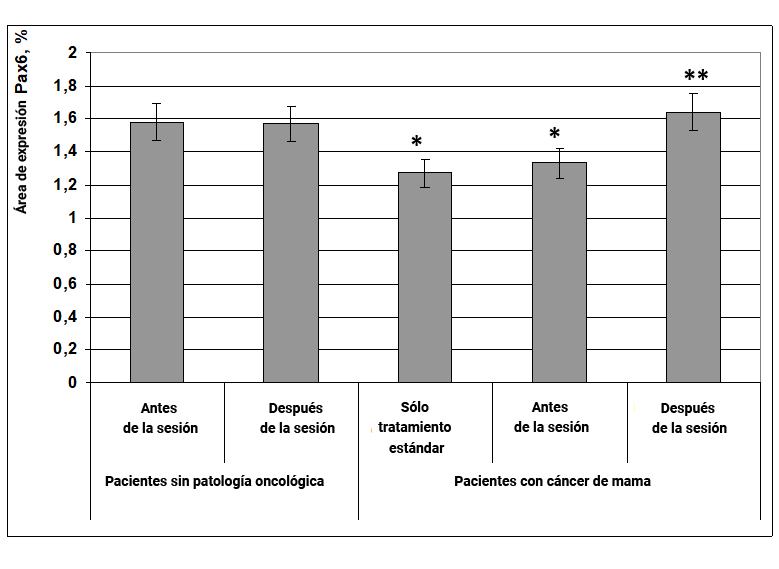
Fig. 7. Influencia del efecto informativo energético sobre la expresión del factor de transcripción Pax6 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
Por lo tanto, el efecto informativo energético contribuye a la normalización de la expresión de la diferenciación del factor de transcripción de las células polipéptidas Pax6 en pacientes con cáncer de mama.
1.2.2. Expresión del factor de transcripción Chx10
Chx10 es un factor de transcripción que regula la diferenciación celular y asegura la formación de una población de células altamente diferenciadas: pinealocitos, fotorreceptores retinianos, miocardiocitos y células epiteliales tímicas, capaces de producir y recibir adecuadamente la melatonina, la serotonina, la proteína pCREB (esas moléculas de señalización que determinan los biorritmos del funcionamiento del tejido y órganos).
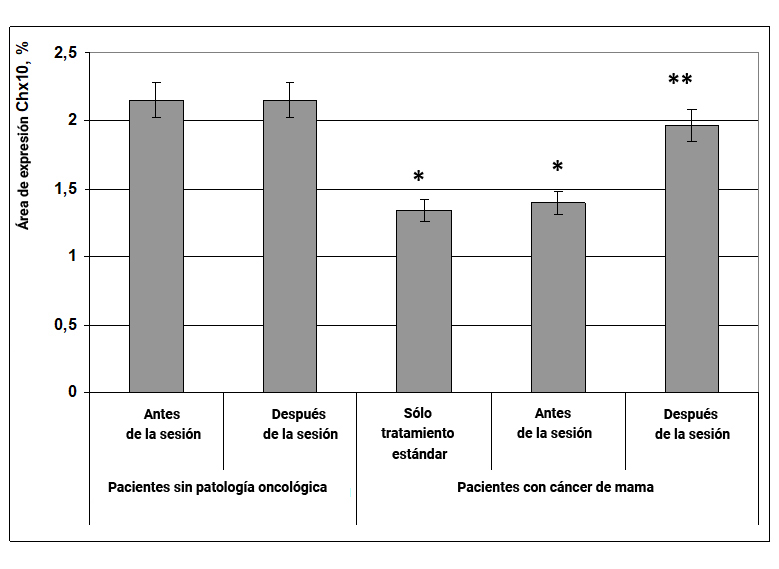
Fig. 8. Influencia del efecto informativo energético sobre la expresión del factor de transcripción Chx10 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
Se demostró que en mujeres sin patología oncológica en el material bucal, el área de expresión del factor de transcripción Chx10 fue casi 2 veces mayor que en pacientes con cáncer de mama (Fig. 8). Al mismo tiempo, el efecto informativo energético en pacientes con cáncer condujo a un aumento significativo en el área de expresión de Chx10 a un nivel normal (Fig. 8, 9).
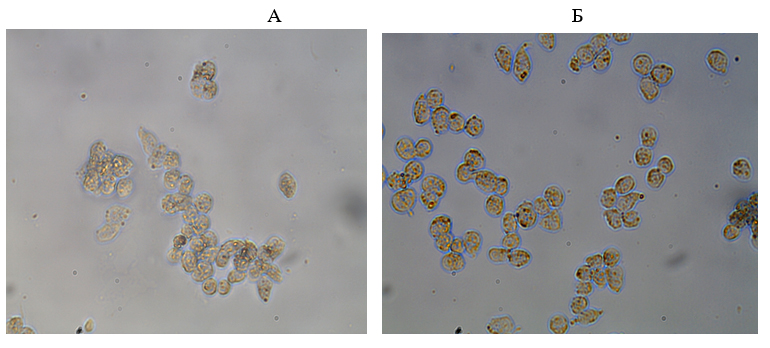
Fig. 9. Expresión del marcador Chx10 en pacientes con cáncer de mama: A - antes de la información y la exposición a la energía, B - después de la información y la exposición a la energía, inmunohistoquímica, x200.
Cabe señalar que una disminución en la expresión del factor de transcripción Chx10 puede ser un factor desfavorable en el curso de enfermedades oncológicas, lo que indica procesos de alteración en la diferenciación y los ritmos circadianos del cuerpo, y el efecto informativo energético normaliza este indicador, teniendo una influencia protectora sobre la diferenciación de las células.
1.2.3 Expresión del factor de transcripción RTF1
El factor de transcripción RTF1 juega un papel importante en la regulación local de los procesos de reparación al proporcionar la función transcripcional de los ribosomas y activar la expresión de genes que codifican un grupo de proteínas de choque térmico involucradas en la termorregulación y la adaptación del cuerpo a diversos efectos estresantes.
Se encontró que en pacientes sin patología oncológica después de la exposición a la energía de la información, el área de expresión del factor de transcripción RTF1 aumentó en un 20% en comparación con el mismo indicador antes de la exposición (Fig. 10). En pacientes con cáncer de mama en los grupos de terapia estándar y antes de la exposición a la información y la energía, el área de expresión de RTF1 fue 1.5 veces menor en comparación con el grupo de mujeres sanas antes de la exposición (Fig. 10).
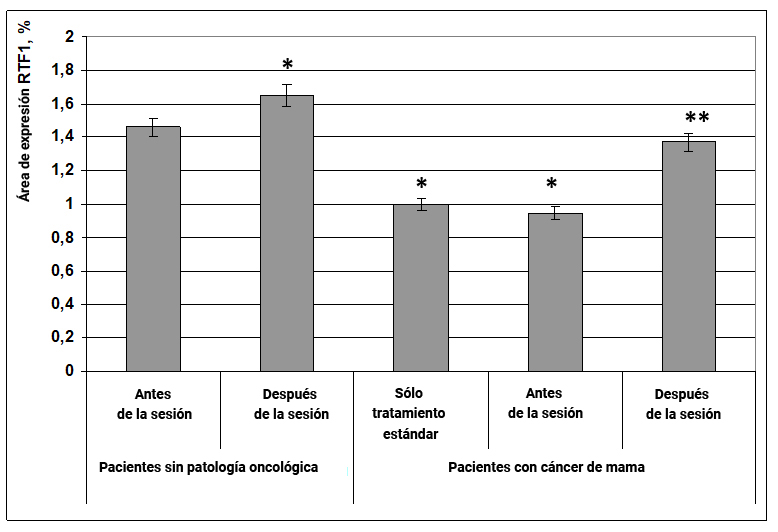
Fig. 10. Influencia del efecto informativo energético sobre la expresión del factor de transcripción RTF1 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
Durante la exposición al efecto informativo energético, el área de expresión del marcador RTF1 en pacientes con cáncer de mama aumentó en un 38% y alcanzó un indicador propio para mujeres sin patología oncológica (Fig. 10).
Los datos obtenidos nos permiten concluir que el efecto informativo energético aumenta la resistencia del cuerpo a los efectos estresantes tanto en pacientes con cáncer como en mujeres sanas.
1.2.4. Expresión del factor de transcripción FOXA2
Es un factor clave en la diferenciación de hepatocitos y otras células epiteliales (por ejemplo, células acinares del páncreas, epiteliocitos de la vesícula biliar).
Por lo tanto, FOXA2 induce la síntesis de albúmina, transtiretina, enzimas pancreáticas y biliares por los hepatocitos, lo cual es importante para la regulación del metabolismo y el desarrollo del páncreas, el hígado y el sistema biliar.
Descubrimos que la expresión del factor de transcripción FOXA2 no cambia con la aparición de cáncer de mama y, por lo tanto, este marcador molecular no es informativo para evaluar el mecanismo de acción del efecto informativo energético en el cuerpo humano.
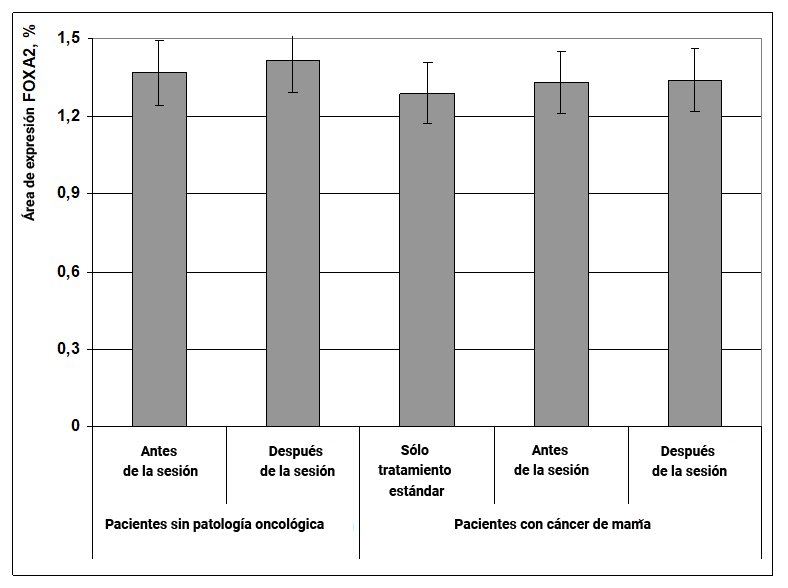
Fig. 11. Influencia del efecto informativo energético sobre la expresión del factor de transcripción FOXA2 en pacientes con cáncer de mama.
1.2.5. Expresión del factor de transcripción Oct2
El factor de transcripción Oct2 juega un papel importante en la regulación de la diferenciación de varias subpoblaciones de linfocitos. Esta molécula de señalización regula la expresión de genes de inmunoglobulina. Induce la producción de moléculas biológicamente activas con actividad citotóxica que contribuyen a la eliminación de aminas endógenas o xenobióticos, incluidas toxinas ambientales, aminas neurotóxicas y fármacos.
En pacientes sin patología oncológica, el efecto energético de la información no afectó la expresión del factor de diferenciación de células inmunes Oct2, pero en pacientes con cáncer de mama este efecto causó un efecto significativo. Por lo tanto, en pacientes con neoplasias, el área de expresión del marcador Oct2 en comparación con mujeres sanas disminuyó en un 25% (Fig. 12). Bajo la influencia del campo de información-energía, el área de expresión de Oct2 aumentó en un 35% (Fig. 12) y alcanzó los valores típicos para mujeres sanas.
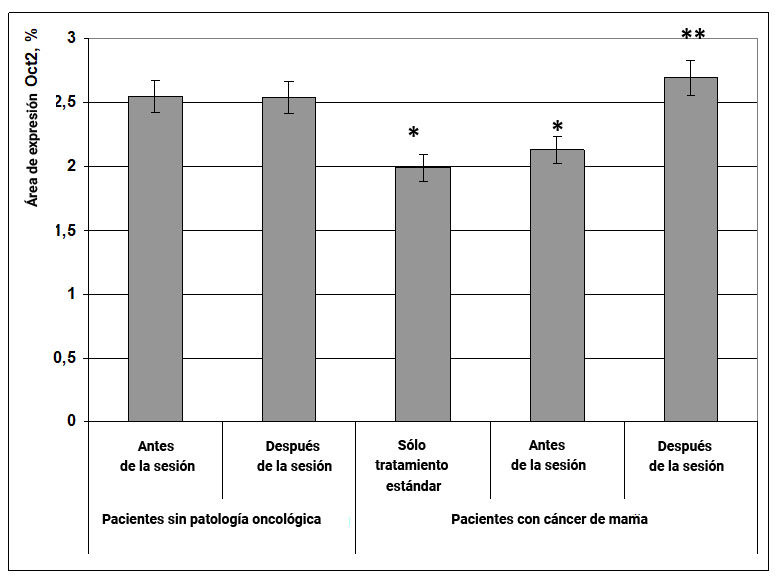
1.2.6. Expresión del factor de transcripción Prox1
El factor de transcripción Prox1 es un regulador específico y necesario del desarrollo de los sistemas circulatorio y linfático.
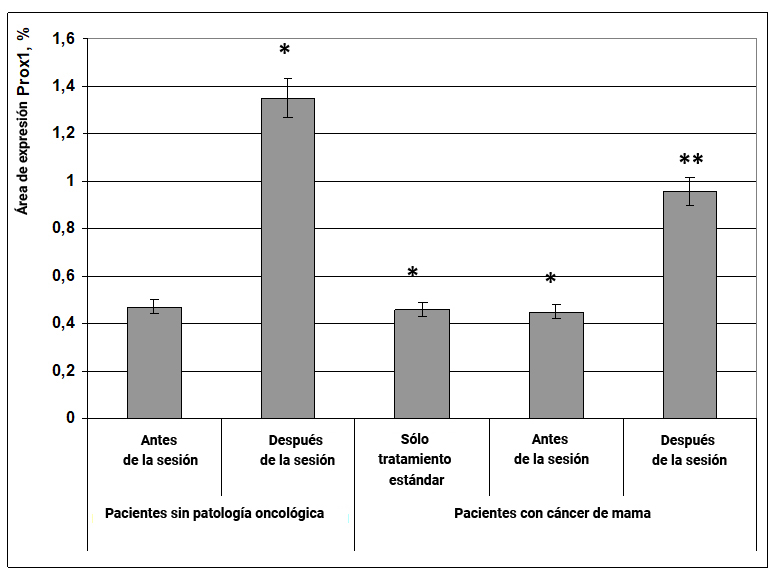
Fig. 13. Influencia del efecto informativo energético sobre la expresión del factor de transcripción Prox1 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
Es la expresión de Prox1 que asegura la maduración adecuada de las células comprometidas (células progenitoras) en formas diferenciadas de células endoteliales. Además, Prox1 proporciona la diferenciación de células comprometidas en tres tipos de células retinianas: horizontal, bipolar y amacrina. Por lo tanto, la función de Prox1 es un factor necesario y clave que garantiza la diferenciación y el mantenimiento oportunos de un grupo de células altamente diferenciadas funcionalmente activas.
El efecto informativo energético ejerció un impacto extremadamente pronunciado sobre la expresión del marcador de diferenciación celular del sistema circulatorio y linfático prox1, tanto en mujeres sanas como en pacientes con cáncer de mama. En mujeres sin patología oncológica, bajo la influencia del efecto informativo energético, el área de expresión de Prox1 aumentó 3 veces (Fig. 13), mientras que en pacientes con cáncer de mama después de la sesión, la expresión inicialmente reducida de este marcador aumentó a valores superiores a los obtenidos para grupos de mujeres sanas (Fig. 13).
1.2.7. Expresión del factor de transcripción Nkx2.5
El factor de transcripción Nkx2.5 desempeña un papel importante para garantizar la actividad de los genes que determinan la diferenciación específica de las células de los tejidos, especialmente aquellas involucradas en el desarrollo y funcionamiento del corazón y los vasos sanguíneos. La mutación en el gen que codifica la expresión de la proteína Nkx2.5 conlleva anormalidades en el desarrollo embrionario del corazón hasta la pérdida completa del marcador de este órgano. La sobreexpresión de este factor durante el desarrollo embrionario y postnatal temprano conduce a la formación de defectos cardíacos, mientras que una disminución en su expresión en el organismo adulto se acompaña por el desarrollo de alteraciones del ritmo, ataques cardíacos e insuficiencia cardíaca.
Se encontró que el efecto informativo energético aumenta la expresión del factor de transcripción Nkx2.5 en mujeres sin cáncer en un 25% (Fig. 14), lo que indica su efecto protector sobre las funciones del sistema cardiovascular.
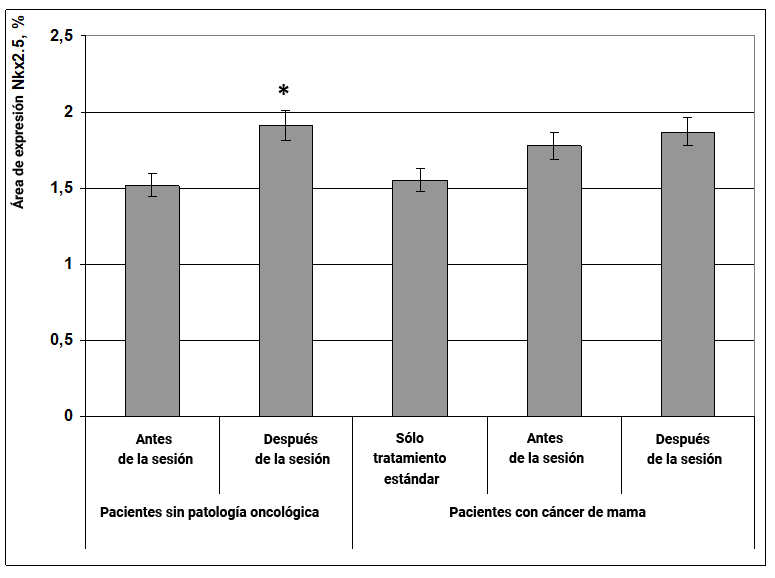
Fig. 14. Influencia del efecto informativo energético sobre la expresión del factor de transcripción Nkx2.5 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
Además, en el cáncer de mama, la expresión de este marcador en comparación con mujeres sanas no cambia significativamente (Fig. 14).
1.2.8. Expresión del factor de transcripción MAB045
MAB045, también conocido como PSD-95 (proteína postsináptica-95) o como SAP-90 (proteína-90 asociada a la sinapsis), es un miembro de la familia de guanilato-quinasa unida a la membrana. Este factor de transcripción juega un papel regulador clave en la formación de canales iónicos en la membrana celular.
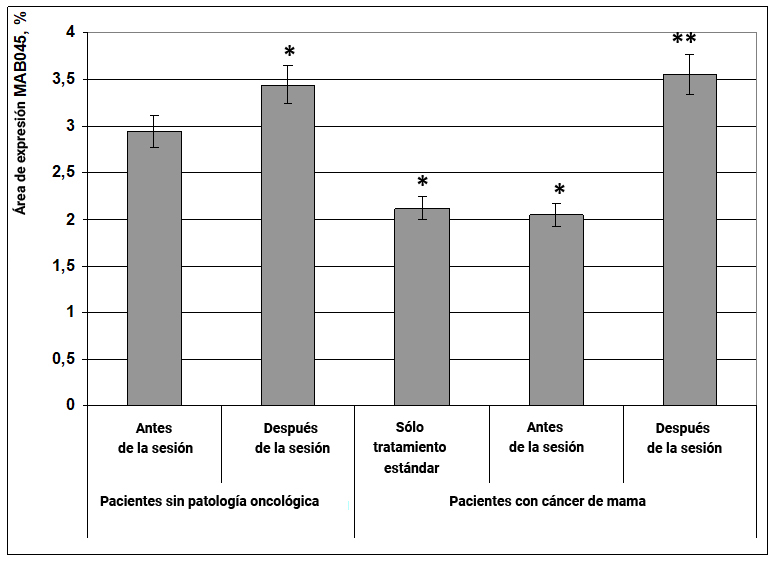
Fig. 15. Influencia del efecto informativo energético sobre la expresión del factor de transcripción MAB045 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
Esta proteína proporciona densidad celular y adhesión de neuronas durante la formación de sinapsis, y también forma complejos de receptores para receptores de glutamato trópico. La disminución de la expresión de este factor conduce a una transmisión sináptica deteriorada, especialmente en los campos neuronales glutamato positivos. Se encontró que en pacientes sin patología oncológica después de la exposición al efecto informativo energético, el área de expresión del factor de transcripción MAB045 aumentó con certitud en un 17% en comparación con el mismo indicador antes de la exposición (Fig. 15). En pacientes con cáncer de mama en los grupos de terapia estándar y antes de que se aplicara la exposición al efecto informativo energético, el área de expresión de MAB045 era en un 43% menor en comparación con el grupo de mujeres sanas antes de la exposición (Fig. 15). Durante la exposición al efecto informativo energético, el área de expresión del marcador MAB045 en pacientes con cáncer de mama aumentó en un 75% y fue mayor que en mujeres sin patología oncológica (Fig. 15).
Los datos obtenidos nos permiten concluir que el efecto informativo energético estimula los procesos de transmisión sináptica en las neuronas cerebrales.
1.2.9. Expresión del factor de transcripción CXCL12
La proteína CXCL12 se refiere a las quimiocinas. Se ha establecido que CXCL12 y su objetivo, el receptor celular CXCR4, juegan un papel importante en la metástasis de las células cancerosas a la médula ósea, los ganglios linfáticos, el hígado y el tejido pulmonar.
Se creó un modelo tridimensional de la interacción de la proteína CXCL12 y el receptor CXCR4, y resultó que 2 moléculas CXCL12 deberían unirse para formar un dímero. Resultó que las propiedades del dímero son diferentes de la forma habitual de la proteína: la proteína CXCL12 regular estimula el movimiento celular, mientras que el dímero evita la migración de todo tipo de células. Se descubrió que el dímero CXCL12 podía inhibir la invasión de células cancerosas y la metástasis.
El efecto energético de la información ejerció un efecto extremadamente pronunciado sobre la expresión de la quimiocina anticancerígena CXCL12, tanto en mujeres sanas como en pacientes con cáncer de mama. En mujeres sin cáncer, bajo la influencia de la información y los efectos energéticos, el área de expresión de CXCL12 aumentó en un 41% (Fig. 16), mientras que en pacientes con cáncer de mama después de la sesión, la expresión de este marcador se redujo inicialmente en más de 2 veces a los valores obtenidos para un grupo de mujeres sanas (Fig. 16).
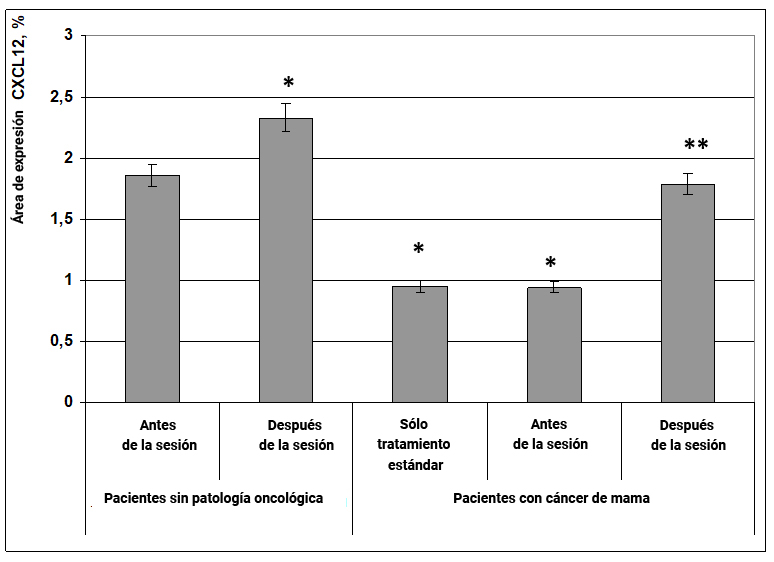
Fig. 16. Influencia del efecto informativo energético sobre la expresión del factor de transcripción CXCL12 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin cáncer antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
Por lo tanto, la influencia terapéutica positiva del efecto informativo energético en pacientes con cáncer de mama se basa en su capacidad para activar la expresión de la proteína anticancerígena CXCL12.
1.2.10. Expresión del factor de transcripción MITF
El factor de transcripción MITF controla el desarrollo y el funcionamiento de las células formadoras de pigmento, en particular los melanocitos.
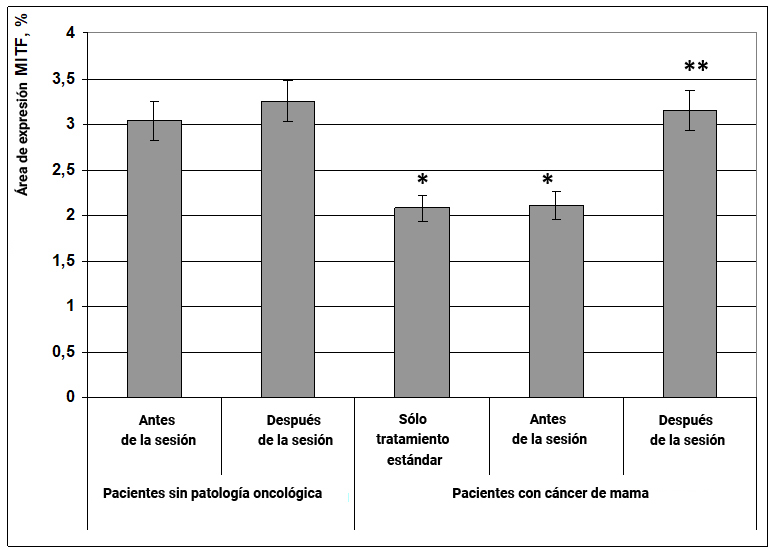
Fig. 17. Influencia del efecto informativo energético sobre la expresión del factor de transcripción MITF en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
Por lo tanto, la expresión de esta proteína proporciona regulación de señalización de la formación de melanina (color de ojos, cabello y piel). Los melanocitos también se encuentran en el oído interno y juegan un papel importante en la función auditiva.
Además, el factor de transcripción MITF está involucrado en la formación del epitelio pigmentario de la retina y una disminución en su expresión implica una reducción de la retina y una percepción alterada de la luz y el color, esto explica la abreviatura MITF, microphthalmia-associated transcription factor, un factor de transcripción asociado con la micro oftalmia. Hay evidencia de la participación de este factor en el desarrollo y diferenciación de osteoclastos y mastocitos inmunocompetentes.
En pacientes sin patología oncológica, el efecto informativo energético no afectó la expresión del factor de diferenciación de las células inmunes MITF, sin embargo, en pacientes con cáncer de mama, este efecto causó un impacto significativo. Por lo tanto, en pacientes con neoplasias, el área de expresión del marcador MITF disminuyó en un 50% en comparación con las mujeres sanas (Fig. 17). Bajo la influencia del campo energético de información, el área de expresión de MITF aumentó en un 54% (Fig. 17) y alcanzó los valores característicos de las mujeres sanas.
Es probable que el efecto informativo energético promueva la activación de las células del sistema inmunitario (mastocitos) a través de la activación del factor MITF, que determina el efecto oncoestático de las sesiones.
1.2.11. Expresión del factor de transcripción BMP2
Se ha establecido que varias proteínas de la familia BMP (proteínas morfogenéticas óseas) se expresan en las células madre neuronales del cerebro de los mamíferos. Además, los factores de transcripción BMP2 y BMP4 tienen efectos supresores sobre las células tumorales. Mediante el análisis cuantitativo de PCR en el cerebro de los mamíferos, se verificaron todas las proteínas de la familia BMP: BMP1, BMP2, BMP3, BMP4, BMP5, BMP6, BMP7 y BMP8a. Es interesante notar que diferentes tipos de células nerviosas expresan la misma cantidad de factores de transcripción BMP2 y BMP4.
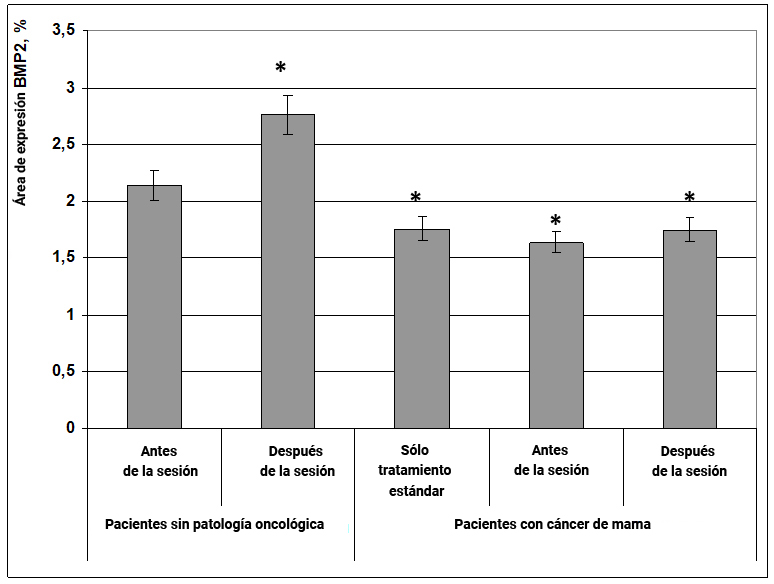
Fig. 18. Influencia del efecto informativo energético sobre la expresión del factor de transcripción BMP2 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
Se encontró que en mujeres sanas, después del efecto informativo energético, el área de expresión del factor de transcripción BMP2 aumentó con certitud en un 27% (Fig. 18). Además, en pacientes con cáncer de mama, el área de expresión del marcador BMP2 se redujo ligeramente, en un 22% en comparación con las mujeres sin cáncer (Fig. 18), lo que puede explicar la ausencia de una influencia significativa del efecto informativo energético en la expresión de este marcador en pacientes con cáncer de mama.
1.2.12. Expresión del factor de transcripción RON
El factor de transcripción RON activa las vías de señalización intracelular (desde la membrana hasta el núcleo celular), proporcionando interacción molecular entre los organóides y el núcleo, es decir. controla la homeostasis intracelular. Una disminución en su expresión conduce a una función deteriorada de las mitocondrias y los ribosomas y puede contribuir a la transformación tumoral intracelular, que se muestra en el material de los tumores de vejiga.
El efecto informativo energético tuvo un pronunciado impacto estimulante sobre la expresión del factor de transcripción RON, tanto en mujeres sanas como en pacientes con cáncer de mama. En mujeres sin cáncer, bajo la influencia del efecto informativo energético el área de expresión de RON aumentó en un 30% (Fig. 19), mientras que en pacientes con cáncer de mama después de la sesión, la expresión de este marcador inicialmente disminuyó en un 36% y aumentó a los valores obtenidos para grupos de mujeres sanas (Fig. 19).
Por lo tanto, el efecto informativo energético tiene un impacto anticancerogénico en el cuerpo al aumentar la expresión del factor antitumoral RON tanto en mujeres sin cáncer como en pacientes con cáncer de mama.
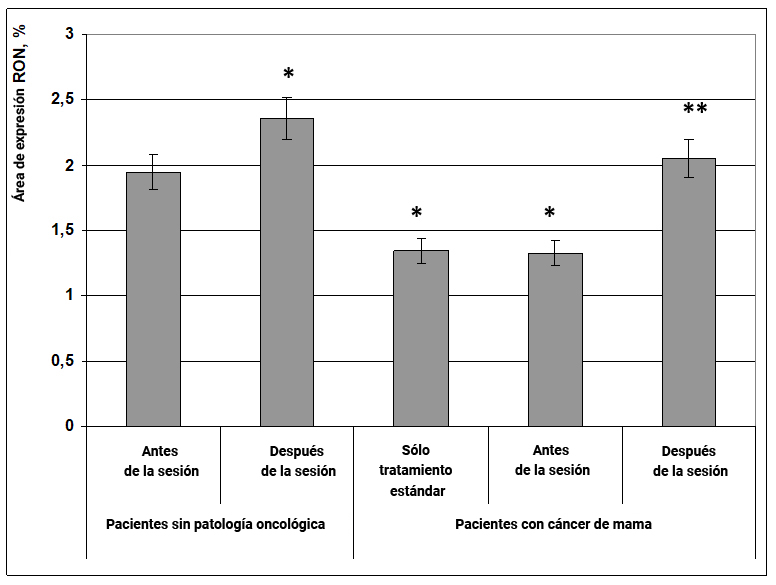
Fig. 19. Influencia del efecto informativo energético sobre la expresión del factor de transcripción RON en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
1.2.13. Expresión del factor de transcripción WNT5A
El factor de transcripción WNT5A es miembro de una gran familia de proteínas cuya síntesis está codificada por el gen WNT5A. Esta proteína está involucrada en la regulación de la renovación celular y, por lo tanto, una disminución en su expresión es uno de los factores desencadenantes de la oncogénesis y estimula la migración celular, lo que es desfavorable para la metástasis. Por el contrario, la mejora de su expresión durante el tratamiento de neoplasias (especialmente melanomas) reduce la proliferación, invasión y migración de células tumorales.
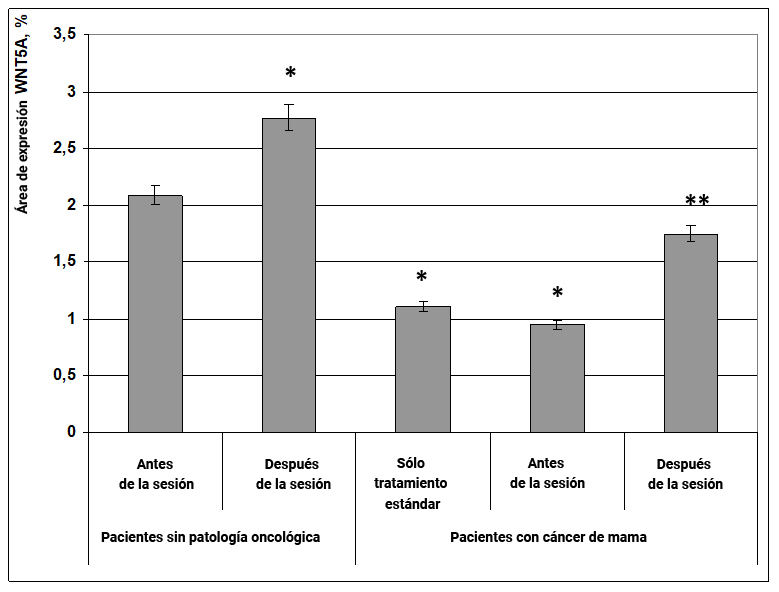
Fig. 20. Influencia del efecto informativo energético sobre la expresión del factor de transcripción WNT5A en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
Nuestros estudios encontraron que el factor de transcripción WNT5A es uno de los principales objetivos del efecto regulador de la información y la exposición a la energía. Entonces, en mujeres sin cáncer después de la sesión, se observó un aumento significativo en la expresión de WNT5A en un 23% (Fig. 20). Al mismo tiempo, en pacientes con cáncer de mama, la expresión de WNT5A disminuyó casi 2 veces en comparación con mujeres sanas, y el efecto informativo energético contribuyó a un aumento significativo en la expresión de esta proteína transcripcional en un 60%, al nivel de control (Fig.20).
Por lo tanto, el efecto informativo energético contribuye a un aumento en la expresión del factor de transcripción WNT5A en mujeres sanas y pacientes con cáncer de mama, lo que indica su efecto antitumoral y una disminución en la probabilidad de metástasis de neoplasias.
1.2.14. Expresión del factor de transcripción SMAD2
El grupo de proteínas SMAD es un mediador citoplasmático activado por interacción con el receptor del factor de crecimiento TGF-beta mediante la fosforilación. Existen varios tipos de SMAD que interactúan con un receptor activado: SMAD1, SMAD2, SMAD3 y SMAD5.
El factor de transcripción SMAD2 media la cascada intracelular de señalización que regula el ciclo celular. Entonces, con su participación, se induce la síntesis de tales inhibidores de la proteína quinasa dependientes de ciclina como p15 y p21. Por lo tanto, el factor de transcripción SMAD2 es una proteína que regula el crecimiento celular y la apoptosis.
Se demostró que en pacientes con cáncer de mama, la expresión de la proteína SMAD2 se reduce ligeramente, y el efecto informativo energético aumenta ligeramente la expresión de esta proteína (Fig. 21), sin embargo, este efecto no es estadísticamente significativo y solo podemos hablar de la tendencia a normalizar este indicador en pacientes con enfermedades oncológicas.
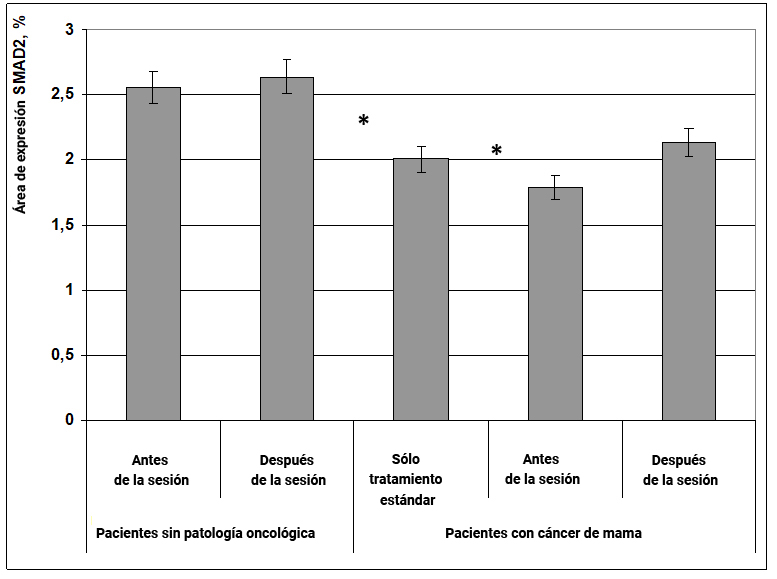
Fig. 21. Influencia del efecto informativo energético sobre la expresión del factor de transcripción SMAD2 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
1.2.15. Expresión del factor de transcripción ERBB3
El factor de transcripción ERBB3 es un receptor de tirosina quinasa y está involucrado en la activación de las vías de señalización intracelular que conducen a la proliferación y diferenciación celular.
Se ha verificado la sobreexpresión del factor ERBB3 para el cáncer de próstata, vejiga y mama.
El efecto informativo energético contribuyó a una disminución en la expresión del pro-oncógeno ERBB3 en mujeres sanas, pero se obtuvo un efecto particularmente pronunciado en pacientes con cáncer de mama. Después de la exposición en mujeres sanas, el área de expresión de ERBB3 disminuyó en un 29% (Fig. 22)
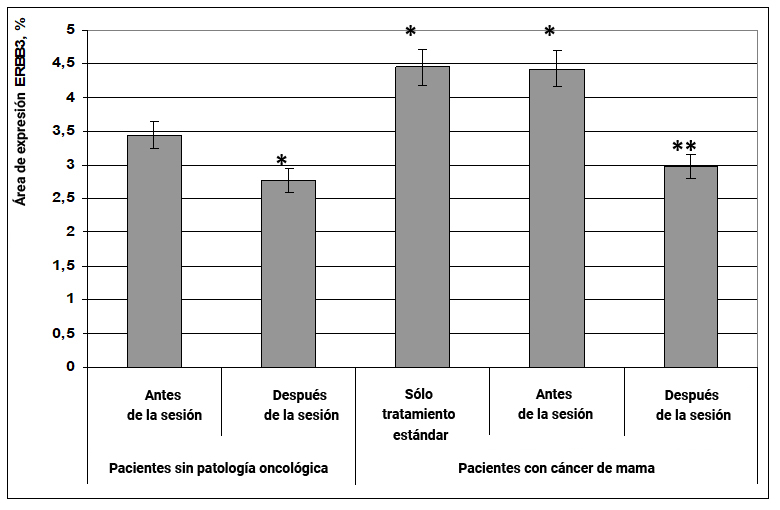
Fig. 22. Influencia del efecto informativo energético sobre la expresión del factor de transcripción ERBB3 en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron solo tratamiento estándar.
Al mismo tiempo, en pacientes con cáncer de mama, se observó la sobreexpresión del factor de transcripción ERBB3, y después de la exposición al efecto informativo energético, el área de expresión de esta proteína disminuyó 1,5 veces (Fig. 22) y alcanzó valores normales característicos de pacientes sin patología oncológica.
1.3. El efecto del campo de información sobre la expresión del péptido mensajero grelina
en pacientes con cáncer de mama
Grelina (G - growth hormone, hormona de crecimiento + relin - un término utilizado en la farmacopea de EE. UU. para referirse a medicamentos que actúan de manera similar a los factores de liberación).
La grelina es un péptido de 28 aminoácidos que circula en la sangre, donde ingresa principalmente como resultado de la secreción del estómago. En las células endocrinas del estómago, la grelina está contenida en gránulos secretores junto con cromogranina A, serotonina y somatostatina.
La grelina se descubrió como resultado de la búsqueda de un ligando endógeno para receptores, a través del cual actúan los derivados sintéticos de meta-encefalina, estimulando la secreción de la hormona del crecimiento por la glándula pituitaria.
La estructura de la grelina resultó ser un péptido similar a la motilina, que es liberada por las células endocrinas del intestino al ingerir alimentos. Las fuentes de grelina son el estómago, los intestinos, los riñones, los huesos y los cartílagos, y la placenta. La grelina también se encuentra en la hipófisis y el hipotálamo. Las células que contienen grelina se encuentran en la tráquea y el esófago. El ARNm para la grelina se encuentra en linfocitos y neutrófilos.
La grelina también se produce en el núcleo arqueado del hipotálamo, donde estimula la secreción de la hormona del crecimiento producida por la glándula pituitaria anterior. Los receptores de grelina se expresan por neuronas en el núcleo arqueado y el hipotálamo ventromedial. El receptor de grelina es un receptor unido a la proteína G (GPCR), anteriormente conocido como el receptor GHS (receptor estimulante de la secreción de la hormona del crecimiento). La grelina desempeña un papel importante en el sistema nervioso, especialmente en el hipocampo, y es importante para la adaptación cognitiva en condiciones ambientales cambiantes y el proceso de alimentación. Recientemente se ha demostrado que la grelina promueve la activación de la isoforma endotelial de la óxido nítrico sintasa en la cascada de señalización, que depende de varias quinasas, incluida la hormona adenocorticotrópica.
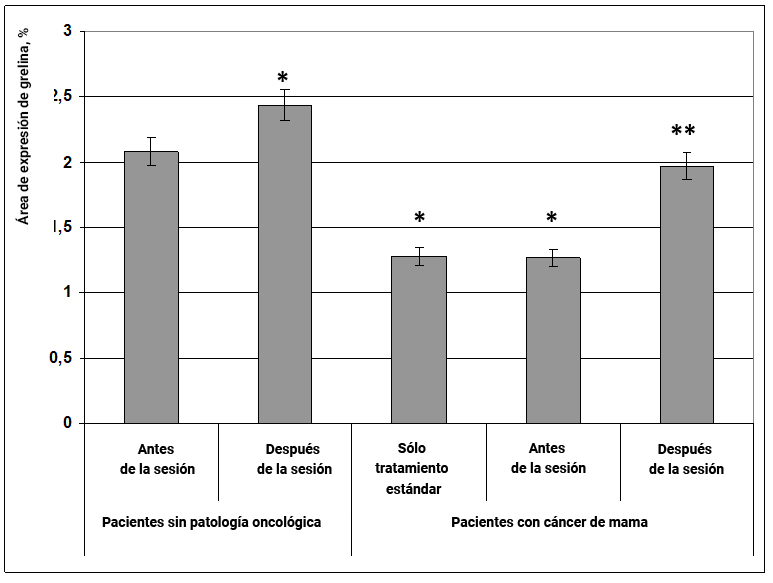
Fig. 23. Influencia del efecto informativo energético sobre la expresión del péptido mensajero grelina en pacientes con cáncer de mama.
* - p <0.05 - comparado con pacientes sin patología oncológica antes de la sesión.
** - p <0.05 - comparado con pacientes que recibieron sólo tratamiento estándar.
Además de los receptores de grelina con los que se encontró, existe otro tipo de receptor, el primero que actúa a través de proteínas G que activan el ciclo de fosfatidilinositol, y el último, probablemente a través de proteínas G que modulan los canales iónicos. Los genes receptores de grelina, como los genes de la grelina en sí, se expresan en linfocitos y neutrófilos.
Como hormona de origen gástrico, la grelina, que actúa sobre el hipotálamo, participa en la regulación del comportamiento alimentario, en particular, estimulando la secreción de la hormona del crecimiento. En el tracto gastrointestinal, la grelina está involucrada en la regulación de la motilidad intestinal.
La información y el efecto energético tuvieron un pronunciado efecto estimulante sobre la expresión del péptido mensajero grelina tanto en mujeres sanas como en pacientes con cáncer de mama. En mujeres sin cáncer, bajo la influencia del efecto informativo energético, el área de expresión de grelina aumentó con certitud en un 10% (Fig. 23), mientras que en pacientes con cáncer de mama después de la sesión, la expresión de este marcador, reducida inicialmente en un 57%, aumentó a los valores obtenidos para un grupo de mujeres sanas (Fig. 23).
Por lo tanto, el efecto informativo energético ejerce una influencia estimulante en la producción de la hormona peptídica grelina, que tiene varios efectos reguladores en el tracto gastrointestinal y el cerebro.
Conclusión
Los datos obtenidos indican que los mecanismos de las células moleculares subyacen al efecto positivo del campo de información (CI) en pacientes con cáncer de mama.
La permanencia de las células en un campo de información altamente concentrado estimula la inmunidad celular, cuya disminución es la razón del desarrollo de tumores, que se expresa para mejorar la expresión de las moléculas CD51 y CD64. Además, después de las sesiones, hubo un aumento en la expresión del marcador de macrófagos CD105 y una mayor síntesis de la glucoproteína CD90, que es responsable de la diferenciación normal de los linfoblastos y previene el desarrollo de leucemia.
Otro mecanismo molecular importante de los efectos creados por el campo de información es su influencia sobre los factores de transcripción. Por lo tanto, la presencia de células en un campo de información altamente concentrado conduce a la normalización de los procesos de diferenciación de varios tipos de subpoblaciones celulares, que se expresa en la estimulación de la expresión de los factores de transcripción Prox1, Oct2, Chx10, Pax6, MITF. Además, la presencia de células en un campo de información altamente concentrado aumenta la resistencia del cuerpo al estrés, lo que se correlaciona con un aumento en la expresión del factor de transcripción RTF1. Cabe señalar que el campo de información contribuye a la normalización de las funciones del sistema nervioso (inducción de la expresión de la proteína MAB045) y del sistema cardiovascular (factor de transcripción Nkx2.5). El campo de información altamente concentrado también mejoró la expresión de grelina, un mensajero de señales peptídicas que apoya la homeostasis del tracto gastrointestinal y el cerebro.
Por lo tanto, el efecto anticancerígeno del campo de información en pacientes con cáncer de mama se basa en mecanismos moleculares celulares que regulan la actividad de los órganos y sistemas más importantes: el sistema inmunitario, el nervioso y el cardiovascular.